|
|
IDENTIDADE PESSOAL:
Caminhos e Perspectivas
Francisco Teixeira (coordenação)
Coimbra, Editora Quarteto, 2004 |
|
AUTOPOIESIS E IDENTIDADE PESSOAL
Francisco Teixeira |
DO SI MESMO BIOLÓGICO AO SI MESMO HUMANO
1. Conduta e sistema nervoso |
No “A Árvore do Conhecimento”, Maturana e Varela descrevem-nos uma experiência com um sapo, realizada em 1945 por J.R.W. Sperry, tendo em vista evidenciar o determinismo estrutural dos sistemas nervosos animais que, por serem biológicos, se encontram encastrados na história das suas acções biológicas e corporais, bem de acordo com a sua máxima segundo a qual “não vemos o ‘espaço’ do mundo mas vivemos o nosso campo visual; não vemos as ‘cores’ do mundo, vivemos o nosso espaço cromático”.
Nos termos daquela experiência, pegou-se num girino (a larva de um sapo) e cortou-se-lhe uma pálpebra (sem lesionar o nervo óptico), de modo a poder rodar-se-lhe o olho 180 graus, após o que se deixou crescer o animal até à idade adulta. Completado o seu processo de amadurecimento pôs-se o sapo operado perante uma mosca, tapando-se o seu olho alterado. Sem hesitações, o sapo lança a sua língua para o insecto, conseguindo capturá-lo de modo perfeito. Após isto repete-se a experiência mas, desta vez, tapa-se não o olho operado mas o olho são, obrigando o sapo a operar em função daquilo que lhe é permitido pelo olho alterado. E o que se observa neste caso é que o animal erra o alvo exactamente em 180 graus. O que se passa é que, se o insecto está à sua frente e em baixo o sapo “dispara” a sua língua exactamente para o lugar inverso, com um desvio, portanto, de 180 graus, ou seja, para trás e em cima. Os resultados da experiência, segundo o relato de Maturana e Varela, são invariáveis, ou seja, são sempre exactamente os mesmos e de acordo com o desvio que o experimentador imprimiu ao olho do animal ainda na sua fase de insipiente desenvolvimento. O que se passa é que o sapo lança a sua língua em função da imagem que foi formada na sua retina, contando, porém, que esta está na sua posição normal e não invertida, como de facto está.
A conclusão que daqui se pretende tirar, e depois de percorridas as páginas anteriores, é quase óbvia. Para o sapo, ao contrário do que supõe o Realismo, não existe um “em baixo” e um “em cima” absolutos, um atrás e um à frente absolutos, num mundo exterior, o mesmo se passando aliás, como nos advertem Maturana e Varela, para os observadores desta experiência. A única coisa de que se pode falar é de um determinado tipo de relações internas ao sistema cognitivo do sapo que prescrevem que, em presença de certa perturbação retiniana, sejam despoletadas certas contracções musculares capazes de moverem a língua, a boca, o pescoço e, no limite, todo o corpo do sapo, em ordem à eliminação dessa perturbação. O que se passa no caso do sapo alterado é que o seu cérebro, em presença de uma perturbação retiniana, opera não através de uma computação de um mundo externo a si mesmo (operação que o conduziria a uma correcção das condições que lhe permitissem caçar o insecto) mas através de uma activação de certas recorrências sensório-motoras entre a posição da retina e certos movimentos musculares, nomeadamente da língua, da boca e do pescoço, ainda que a primeira hipótese pareça a mais evidente a um observador de senso-comum representacionista. O que aqui se pretende destacar, numa primeira abordagem, é que o funcionamento do sistema nervoso, na medida em que este é uma unidade autopoiética de segundo grau (que acopla, portanto, unidades autopoiéticas de primeiro grau), funciona segundo os princípios da determinação estrutural e da clausura operacional, pelo que a conduta daí resultante é uma função das suas relações estruturais internas, ou seja, da sua estrutura de conexões neuronais. A constituição do sistema nervoso diz-nos que quaisquer que sejam as suas mudanças, estas acabam por produzir outras mudanças no interior do próprio sistema com o objectivo de manter constantes certas relações entre os seus componentes particulares. O meio, como já vimos relativamente às unidades autopoiéticas de primeiro grau, não pode determinar o funcionamento do sistema nervoso, mas simplesmente despoletar certo tipo de operações determinadas pela sua estrutura em ordem à sua manutenção enquanto unidade autopoiética de segunda ordem. Ora, acontece que as reorganizações estruturais dos sistemas nervosos dependem das suas histórias particulares de acoplamento estrutural, histórias essas que definem o percurso individual de cada sistema autopoiético desde uma estrutura inicial até uma complexidade crescente de estruturas e a uma capacidade crescente de resistir às perturbações mais diversas. E estas interacções (dadas nos sucessivos acoplamentos estruturais), não será demais dizê-lo, não são interacções instrutivas do meio no sistema (para o qual essa diferença não faz sentido) mas antes perturbações que, pela sua reiteração, tornar-se-ão decisivas na constituição da história estrutural do sistema. É isso que nos querem dizer Maturana e Varela ao referirem que
“a ontogenia de todo o ser vivo consiste na sua contínua transformação estrutural, num processo que, por um lado, ocorre nele sem interrupção nem da sua identidade nem do seu acoplamento estrutural com o seu meio desde o seu início até à sua desintegração final, e por outro lado, segue um caminho particular seleccionado na sua história de interacções pela sequência de mudanças estruturais que estas gatilharam nele” (Maturana e Varela, 1990: 109).
Deste ponto de vista, portanto, aquilo a que chamamos conduta dos organismos não corresponde a um processo interno determinado pelo sistema nervoso (já que o sistema nervoso não se conduz para intenção alguma fora de si mesmo) mas à descrição que dele é feito por observadores, descrição, por definição, externa à identidade do próprio sistema. O que se quer afirmar é que, ainda que possamos dizer adequadamente que as mudanças estruturais de um sistema determinam o estado do organismo observado, a verdade é que o estado e as mudanças de estado do organismo em relação a um ambiente determinado são sempre e somente orientadas pela necessidade dos organismos manterem a sua autopoiesis, pelo que qualquer mudança, sendo embora de um ponto de vista exterior sempre observada como uma conduta, nada diz quanto à intencionalidade do sistema (embora possa dizer algo, e frequentemente diz, a respeito das expectativas do observador). No entanto, se o funcionamento do sistema nervoso nada nos diz quanto a uma actividade conductual intencional, quer dizer, nada nos diz a respeito de finalidades relativamente às suas interacções ambientais, já, pelo contrário, se pode e deve pensar o sistema nervoso como dotado de um dinamismo de autocorrecções e equilibrações sistemáticas que, de modo dramático, luta permanentemente pelo enrijecimento e pela plasticidade das suas redes celulares de modo a “ampliar o campo das possíveis correlações sensório-motoras do organismo e a expandir o domínio da conduta” (Maturana e Varela, 1990: 140) (1). É o que acontece com certo tipo de células receptoras (para além daquelas outras que têm a finalidade de se oferecer às perturbações externas) que têm a função de induzir mudanças globais no sistema neuronal quando detectam certas anomalias no meio interno em função das quais essas anomalias são corrigidas. Maturana e Varela apresentam-nos o caso específico de certas células quimio-receptoras de determinadas artérias que alteram o seu estado em função da mudança das diferentes concentrações presentes no meio sanguíneo dos vertebrados. Em função das suas mudanças de estado, despoletadas pelas mudanças das concentrações sanguíneas, essas células vão modificar, por sua vez, o estado de certos neurónios que através das suas mudanças contribuem para uma re-equilibração das concentrações sanguíneas, que por sua vez despoletaram todo o processo. Estas mudanças podem ou não dar origem a comportamentos motores diferenciados. Pode então dizer-se, na bela expressão de Maturana e Varela, que “toda a conduta é uma visão externa da dança de relações internas do organismo” (Maturana e Varela, 1990: 141). A conduta de um organismo corresponde, pois, a uma atribuição externa e não a uma finalidade orgânica internamente determinada. Caberá, depois, ao investigador descobrir as coerências ou circuitos neuronais que são responsáveis por esta ou aquela conduta.
No entanto, e naturalmente, as coerências neuronais funcionam segundo certo grau de invariabilidade comum aos membros de determinada espécie. A plasticidade neuronal não é uma plasticidade ilimitada, sendo constrangida por linhas de conectividade que especificam certas arquitecturas comuns. De igual modo, como de alguma forma já foi ficando implícito, as mudanças estruturais dos sistemas neuronais como resposta a perturbações, não são nem podem ser efeito de conexões sinápticas particulares mas sim de padrões de conectividade locais. As fendas sinápticas não correspondem a espaços privativos de relações entre neurónios, mas consistem antes em espaços de múltiplas inter-relações químicas e eléctricas determinadas por outros neurónios, em função de outras fendas sinápticas banhadas por elementos carreados através da corrente sanguínea e provenientes de todas as partes do organismo, segundo concentrações muitas vezes despoletadas por perturbações externas ao sistema. As relações entre os neurónios resultam, então, de delicados equilíbrios, matizados por um elevadíssimo número de elementos que despoletam mudanças locais. Deste ponto de vista, virtualmente todas as relações entre o sistema e outros elementos são susceptíveis de causar mudanças estruturais no sistema, sejam elas mais ou menos evidentes, mais ou menos imediatas ou mais ou menos diferidas nos tempo (2).
Pode dizer-se que esta é uma descrição, embora muito geral, do tipo de funcionamento segundo o qual os “organismos nervosos” alargam os seus domínios de interacções. Trata-se, então, de “acoplar as superfícies sensoriais e motoras mediante uma rede de neurónios cuja configuração pode ser muito variada” (Maturana e Varela, 1990: 136) (3).
Há pois uma plasticidade de altíssimo grau que é intrínseca às arquitecturas neuronais, particularmente às dos mamíferos superiores. Convém que fique claro, porém, que essa plasticidade é relativa não a uma presumível capacidade adaptativa ao meio, através de reordenações neuronais sucessivas em ordem à sua correspondência óptima com um meio sucessivamente internalizado, mas, pelo contrário, é relativa à sua capacidade de sucessivas mudanças estruturais, de forma a manter o seu acoplamento estrutural com o meio em resultado de contínuas perturbações. Maturana e Varela advertem-nos que, do ponto de vista do observador, estas reordenações estruturais da organização sistémica nervosa podem ser, e frequentemente são, entendidas como uma aprendizagem adequada. Do mesmo modo, essas mudanças, podem ser entendidas como correspondendo às necessidades impostas pelo meio. No entanto, isso só é assim de maneira muita ampla, porquanto as condições do meio não especificam positivamente mas só negativamente as derivas orgânicas dos sistemas autopoiéticos. O que acontece verdadeiramente é que o organismo e o meio coexistem num processo de interacção e causação de mudanças estruturais, congruentes com as suas mútuas manutenções de identidade e autonomia. A não ser assim, como também já dissemos, acontece a morte e ou a desintegração.
Uma das consequências desta sucinta descrição do sistema nervoso e dos seus âmbitos conductuais é o reforço de tese central do pensamento construtivista e autopoiético segundo a qual o conhecimento, humano e não humano, não opera através de representações do mundo. O sistema nervoso, e em particular o sistema nervoso humano, funciona com determinismo estrutural e clausura operacional, pelo que o meio não pode determinar a natureza das imagens, dos conceitos e das acções do organismo. O que o meio pode fazer é engatilhar ou despoletar determinadas configurações neuronais correspondentes a essas imagens, conceitos e acções, mas nada nos pode dizer acerca da sua natureza semântica ou sintáctica (4). Acontece que, porque o sistema nervoso funciona com determinismo estrutural e clausura operacional, ele constitui uma rede neuronal fechada para quem a própria corporalidade em que se aloja constitui o seu ambiente, quer dizer, o universo dos seus elementos externos de perturbação. No caso do sistema nervoso, acoplamento ao meio quer dizer acoplamento ao corpo que lhe é próximo, através de células sensoras e efectoras que, porém, também neste caso, não põem em causa a clausura operacional do sistema, uma vez que o que ocorre nelas não é captação ou a transmissão de informação, mas sim a capacidade de se deixar perturbar e de perturbar o seu ambiente (veja-se o conceito de transdução na nota 8).
Maturana e Varela convidam-nos, com o seu pensamento autopoiético, a considerar uma via média entre o solipsismo e o realismo objectivista, uma via que considere a natureza sistémica (e portanto auto-referente) da nossa cognição, mas que também seja capaz de considerar as suas relações com o contexto que o transcende, introduzindo a distinção entre os domínios diferenciados do sistema e do observador como a distinção capaz de conciliar a compreensão do funcionamento circular dos sistemas biológicos com a compreensão das relações desses sistemas com o meio ambiente. No entanto, a compreensão destes dois domínios (os domínios do observador e do sistema) exige que a reflexão epistemológica os mantenha rigorosamente separados, sob perigo de equívocos permanentes e de longo alcance. Feito isto, mantendo rigorosamente separados os domínios respectivos de observação e do sistema, “não necessitamos de recorrer a representações, nem necessitamos de negar que o sistema opera num ambiente com o qual é comensurável como resultado da sua história de acoplamento estrutural” (Maturana e Varela, 1990: 116)(5). Então, tendo em conta a nossa pesquisa daquilo a que poderemos adequadamente chamar conhecimento e tendo assente esta divisão central entre os domínios do observador e do sistema, poderemos dizer, com alguma segurança, que toda a operação de um sistema autopoiético, observada como uma conduta, pode ser entendida como um acto de conhecimento.
Podemos assim retornar, de modo argumentativamente sustentado, à premissa central do paradigma construtivista, particularmente do paradigma autopoiético, segundo o qual viver, enquanto manutenção do acoplamento estrutural de um ser vivo com o seu meio, é conhecer. E, neste processo de sobrevivência através de contínuos acoplamentos estruturais, o sistema nervoso é duplamente decisivo no processo cognoscitivo: através da ampliação dos estados possíveis do organismo, em virtude da sua imensa plasticidade capaz de “configurações sensório-motoras” extremamente diversificadas e também em virtude de permitir ao organismo formas variadas de acoplamentos estruturais com o meio ambiente, que são a chave da sua sobrevivência (Maturana e Varela, 1990: 149). E vai ser exactamente o sistema nervoso e a sua complexidade relativa que vai introduzir no reino do vivo a diferença fundamental entre aquelas espécies que detêm um operar ou um conhecer relativamente limitado, e aquelas outras que podem operar reestruturando-se e acoplando-se de modo quase infinito.
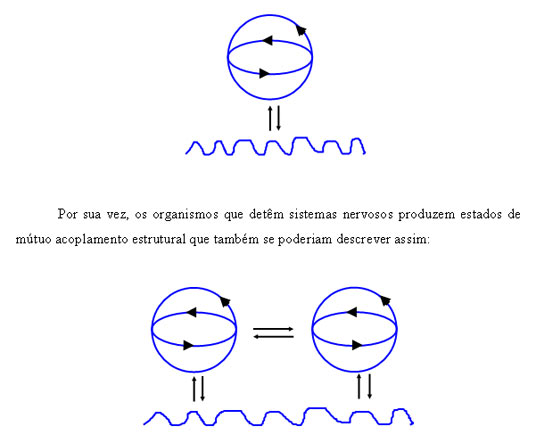
Assim, nos termos das representações gráficas de Maturana e Varela, poderíamos representar os sistemas autopoiéticos dotados de sistemas nervosos do seguinte modo:
|
|
De modo similar à descrição do acoplamento entre organismos multicelulares, o acoplamento entre organismos dotados de sistema nervoso percorre uma lógica comum segundo a qual o outro organismo é entendido como elemento de perturbação, sem nenhuma distinção essencial relativamente ao resto do meio ambiente não vivo. No entanto, o que se espera é que estas perturbações, pela sua recorrência, permitam a constituição de reordenações estruturais congruentes, ou seja, de um acoplamento estrutural capaz de manter a identidade de cada sistema e, portanto, a sua autopoiesis relativa.
Quando ocorrem acoplamentos estruturais através de reordenações estruturais recorrentes entre organismos dotados de sistemas nervosos, Maturana e Varela descrevem este tipo de acoplamento como próprios de uma fenomenologia de terceira ordem. Estes acoplamentos de terceira ordem são decisivos na medida em que, sem eles, não seria possível, por um lado, o tipo de interacções necessárias à reprodução (nomeadamente no caso da reprodução sexuada) e, por outro, o crescimento dos jovens que, num relativamente grande número de espécies, necessita de protecção familiar e de um processo de aprendizagem social sem os quais a sobrevivência não seria possível. Por estas razões os fenómenos de terceira ordem são aqueles que incluem algum grau de sociabilidade.
No homem, porém, a fenomenologia de terceira ordem referida, ou seja, a natureza social das suas relações, embora necessária exactamente pelas mesmas razões aduzidas para o conjunto de outros seres vivos com sistema nervoso, produz resultados que, pela riqueza desses sistemas, lhe permite a produção de certos domínios de acoplamento com outros homens que fazem emergir um conjunto de fenómenos novos característicos da sua espécie e da sua deriva onto e filogenética que, por sua vez, lhe abrem novos e mais amplos domínios de acoplamento. Estamos a falar, naturalmente, da linguagem humana e da sua capacidade auto-reflexiva, do si mesmo e da mente ou, como diria Rorty, da sua capacidade específica de descrições e redescrições sucessivas de si mesmo, aquilo que faz com que, afinal, o homem se distinga de modo decisivo das outras espécies vivas (6). Ou seja, a linguagem humana emerge das coordenações conductuais de terceira ordem através de uma atribuição de valor semântico a essas condutas, valores esses que constituem, autenticamente, o conjunto da nossa vida mental. Acontece que, assim sendo, o si mesmo psíquico (a identidade pessoal ou o self) não é de natureza autopoiética, no sentido em que não é redutível à biologia, já que emerge das relações sociais, ou, mais exactamente, dos acoplamentos estruturais, embora, naturalmente, pressuponha e necessite de um nível de sustentação intrinsecamente autopoiético. Ou seja, o si mesmo psíquico (e já não redutoramente biológico) é algo que emerge através de uma atribuição de valor semântico a uma conduta, quer dizer, é algo que, estando embora encastrado biologicamente, não pode ser reduzido aos sistemas biológicos enquanto unidades autopoiéticas de segunda ou primeira ordem, já que emerge do relacionamento social, que, no caso específico do homem, é um relacionamento social intrinsecamente linguístico.
Sintetizando: para Maturana devem distinguir-se nos sistemas autopoiéticos de segunda ordem (compostos) dois domínios diferenciados e não intersectáveis de descrição: os domínios da fisiologia e da conduta. O domínio da fisiologia diz respeito ao domínio sistémico estrutural, próprio dos vários componentes da unidade; já o domínio da conduta diz respeito ao domínio fenoménico da unidade autopoiética, que é especificado pelas interacções da unidade composta com o ambiente, enquanto unidade simples, quer dizer, percebida e entendida como uma totalidade: é o domínio dos fenómenos percebidos como comportamentais, do ponto de vista do observador externo à estrutura do sistema. Os dois domínios, como já vimos, os domínios da fisiologia e da conduta, ou os pontos de vista do sistema e do observador, não podem ser confundidos, sob perigo de ocorrerem nefastas confusões epistemológicas.
|
| |
(1) A natureza recursiva dos sistemas nervosos ou neuronais (presentes na quase totalidade das derivas filogenéticas) e a sua correspondente força expansiva e plástica é notavelmente demonstrada por Heinz von Foerster num artigo clássico intitulado “Construindo uma Realidade”, ainda que dizendo apenas respeito a certas formas básicas e primordiais deste tipo de fenómenos. Nesse artigo de poucas páginas, von Foerster mostra-nos como os “efectores independentes”, ou unidades sensório-motoras como as que apresentam os protozoários e os metazoários distribuídos pelas suas superfícies externas, podem ter estado na origem dos neurónios e correlativos sistemas neuronais. Estes “efectores independentes” são constituídos por uma secção sensível e outra motora intimamente relacionadas, de modo que um efeito químico sobre a secção sensível tem imediatamente um efeito correlativo de contracção sobre a secção motora. Esta contracção sobre a secção motora pode provocar, e frequentemente provoca, uma mudança de forma ou de movimento do animal, que, por sua vez, pode conduzir a novas interacções químicas com o meio, provocando por sua vez novas contracções com novos movimentos e ou novas formas do animal e assim sucessivamente de modo recursivo e permanente, até à separação das sub-unidades sensoriais das sub-unidades motoras. O que nos diz von Foerster é que este desligamento entre as secções sensorial e motora dos “efectores independentes” levou consigo o desenvolvimento de finos ligamentos (os axónios) que transmitem as perturbações do sensório ao motor, dando origem “ao conceito de sinal: observa algo aqui, actua correspondentemente ali” (Foerster, 1998: 44-45). Mas o passo mais decisivo neste processo de ampliação das correlações sensório-motoras do organismo parece ter estado, segundo von Foerster, no aparecimento de um neurónio interposto entre a unidade sensorial e a unidade motora. Este neurónio, não sendo mais que uma célula sensorial, tem no entanto as estranhas características de, por um lado, funcionar apenas em presença de um agente universal, a actividade eléctrica de axónios aferentes e, por outro, de alterar a sua capacidade de acção presente em virtude da sua carga eléctrica anterior, introduzindo no reino animal o conceito de computação enquanto capacidade de ampliação das formas de conduta. Ora, a partir da rápida multiplicação destes neurónios compreende-se facilmente como se procedeu à sofisticação das conexões horizontais dos sistemas neuronais, dando origem aos cérebros.
(2) Freud certamente ficaria satisfeito com esta asserção do entendimento autopoiético dos sistemas nervosos. É que, ao defendermos que determinado tipo de acoplamentos estruturais entre um sistema nervoso e o meio pode ter efeitos diferidos no tempo em virtude de certos reordenamentos neuronais de natureza local, pode permitir uma aproximação da máxima freudiana segundo a qual todo o tipo de relações humanas e não humanas é susceptível de, de modo indeterminado e diferido no tempo, provocar reordenações mentais conscientes e não conscientes.
(3) Justifica-se aqui uma descrição muita sucinta da estrutura neuronal dos organismos mais complexos como os mamíferos superiores. Nestes organismos todos os neurónios têm funções especializadas. Poderemos começar por dizer que alguns neurónios estão ligados a células receptoras especializadas em responder a diferentes energias externas ao sistema (pressões físicas, presença da luz, alterações químicas, etc...) e, nalguns casos, em transformar essas energias (o termo técnico para esta transformação é transduzir) em impulsos eléctricos ou nervosos reconhecíveis pelo sistema neuronal. Sem este efeito de transdução a energia externa ao sistema (ou perturbação ao sistema) não seria reconhecível ou, então, poderia provocar a sua morte. Esta transdução tem o efeito de levar os impulsos nervosos a produzirem alterações neuronais. Estes neurónios, que enviam os impulsos nervosos das células receptoras para outras configurações neuronais mais complexas do sistema nervoso, têm o nome de neurónios sensoriais. Como acontece com certos neurónios responsáveis pelas sensações tácteis, por vezes o elemento receptor da energia exterior é uma parte especializada destes neurónios sensoriais. No entanto, na maioria das vezes, as funções receptora e de transdução são de responsabilidade celular diferenciada. É o que acontece com a audição e a visão, em que a transdução da estimulação óptica e atmosférica é operada por determinadas células receptoras que, em virtude de certas transformações, produzem certos efeitos em determinados neurónios que, por sua vez, activam outros efeitos neuronais de zonas cerebrais muito diversificadas (no entanto, a visão, sendo o mais estudado dos sistemas perceptivos animais é também, e talvez por isso, o mais rico de interpretações contraditórias). Para além destes neurónios outros há que têm longos axónios que terminam em células efectoras. É o caso dos motoneurónios, que activam a musculatura estriada, ou seja, a musculatura que suporta o esqueleto das pernas e dos braços. A base celular dos motoneurónios está, regra geral, fixada ou na espinal medula ou no cérebro e liga-se directamente através dos axónios com as células efectoras dos músculos. Pode dizer-se que, do ponto de vista autopoiético, quando uma célula receptora é tocada por uma perturbação exterior ao sistema ela produz uma transdução dessa perturbação canalizada através de um neurónio sensorial que, por sua vez, poderá interagir com um motoneurónio que promoverá a libertação de determinada substância química que tem por função a contracção das fibras nervosas provocando um movimento muscular. No entanto, esta resposta tem por finalidade não responder a qualquer informação externa, mas simplesmente manter estáveis os equilíbrios estruturais do sistema. Mas, para além dos sensoriais e dos motoneurónios, a maior parte dos neurónios dos organismos superiores são os interneurónios, ou neurónios situados entre os sensoriais e os motoneurónios. Todos somados, os neurónios do sistema nervoso humano, por exemplo, ascendem a cerca de cem mil milhões, com um elevadíssimo grau de interconectividade entre si mesmos (através das dendrites e das teleodendrites, no primeiro caso extensões do corpo celular através das quais se recebem impulsos nervosos de outros neurónios e, no segundo, terminais do axónio através dos quais se enviam impulsos nervosos a outros neurónios) e com as células receptoras. Supõe-se que os neurónios do cerebelo, por exemplo, especializados na coordenação motora, recebam impulsos de oitenta mil outros neurónios. Ainda que este tipo de neurónios seja aquele que manifesta uma maior interconectividade, pode, descontando-se essa conectividade para um valor médio, imaginar facilmente até que ponto a conectividade neuronal dos mamíferos superiores pode ser fantasticamente elevada (Gleitman, 1993: 22-60).
(4) Como já vimos, a acusação recorrente a este tipo de compreensão sistémica do sistema nervoso e cognitivo é de que ela transporta consigo uma concepção solipsista e idealista da cognição, concepção inviável para a compreensão da nossa acção no mundo porque incapaz de explicar a relação, ou a comensurabilidade (para utilizar a terminologia de Kuhn), entre o homem e o mundo. Estas últimas linhas têm como objectivo de dar a entender como essa acção no mundo é possível, ultrapassando assim as acusações recorrentes daqueles que são incapazes de compreender a cognição para além do senso comum platónico/representacional, obscurecendo e esquecendo a natureza biológica do conhecimento humano e as suas consequências epistemológicas.
(5) Numa das muitas e interessantes gravuras do “A Árvore do Conhecimento”, Maturana e Varela representam esta luta permanente entre o realismo e o solipsismo como o terrível momento da viagem dos argonautas em que estes têm que passar entre o remoinho Caríbdis e o terrível monstro de várias cabeças Cila. Os argonautas são, como se sabe, os viajantes da nave Argos, que transporta o melhor da juventude grega em busca do Velo de Ouro, que representa o espírito de Frixo, filho do rei grego Àtamas, que é salvo dos ímpetos assassinos da segunda mulher de seu pai por um carneiro mágico com um velo de ouro. O Velo de Ouro representa, então, o regresso à justiça dinástica originária e apresenta-se, eventualmente, como uma metáfora da busca da justiça. No entanto, o que eu queria aqui referir é que, embora Maturana e Varela nos apresentem Cila como um monstro de muitas cabeças, o mito original apresenta Cila não como um monstro mas como um rochedo íngreme e sólido, em contraposição a Caríbdes, um vórtice onde o mar se encontra alteroso e “as ondas furiosas se elevam nos ares até tocar o céu” (Hamilton, 1991: 181). Penso que esta descrição é mais adequada àquilo que, no fundo, Maturana e Varela nos querem dizer, a saber, que a opção entre o realismo entre o solipsismo é a opção entre um rochedo íngreme e assustador (mas com fundações seguras) e um vórtice que tudo suga e onde reina o caos e o terror. A proposta de Maturana e Varela, tal como o fizeram com êxito os argonautas, é a de navegar através destas duas ameaças, de modo equilibrado, sem que se possa evitar, embora, uma contínua sensação de responsabilidade e vigilância nesse navegar.
(6) Para Maturana, o humano define-se não pela autoconsciência mas pela linguagem, já que é esta que é capaz de lhe prover a possibilidade de se autodistinguir enquanto se distingue. Assim, para percebermos o que é específico do humano devemos perguntar-nos não pelas vantagens evolutivas, selectivas, da linguagem e da consciência, mas sim pela especificidade do modo de vida dos ancestrais humanos que levou a que a linguagem emergisse e se consolidasse em determinado filo evolutivo, até ao ponto em que se tornou indispensável para que o filo linguajante, o filo humano, que a linguagem continuasse a desenvolver-se. A humanidade ocorre(u), então, enquanto modo de vida específico que permitiu a emergência de si. Ora, esse modo de vida foi, para Maturana, um modo de vida centrado na cooperação, no amor, na sexualidade frontal, na ternura, no cuidado com os descendentes e isto em grupos relativamente pequenos, de sete a oito indivíduos. Este modo de vida (o modo de vida homo) foi, é, centrado num estilo de cooperação social não hierarquizado, ao contrário, por exemplo, do modo de vida antropóide, centrado na hierarquia e na dominação de um macho (Maturana: 2002).
|
AUTOPOIESIS E IDENTIDADE PESSOAL-INDEX
|
|
|
|
|